Tái tổ hợp tương đồng
Tái tổ hợp tương đồng, bao gồm hoán vị gen là một kiểu tái tổ hợp gen trong đó các trình tự nuclêôtit của hai đoạn DNA tương ứng nhau được trao đổi cho nhau.[1], [2]

Thuật ngữ này xuất xứ từ tiếng Anh homologous recombination (tức là tái tổ hợp tương đồng) dùng để chỉ trao đổi các đoạn DNA tương ứng nhau giữa hai nhiễm sắc thể tương đồng.Trong bài này, thuật ngữ tái tổ hợp tương đồng sẽ viết tắt theo tiếng Anh là HR (từ Homologous Recombination). Vì quá trình này diễn ra trong giảm phân của sinh vật nhân thực, nên cũng còn gọi là tái tổ hợp giảm phân (meiotic recombination), hay tái tổ hợp nhân thực (eukaryotic DNA recombination).[3]
Ngoài ra, sau khi phát hiện cả vi khuẩn cũng có HR, thì trường hợp này gọi là tái tổ hợp nhân sơ (Recombination in Prokaryotes). Tuy nhiên ở sinh vật nhân sơ không có cặp nhiễm sắc thể, nên ở nhân sơ thì thực chất hiện tượng này là trao đổi đoạn tương hỗ giữa hai phân tử DNA.[4], [5]
Lược sử nghiên cứu
sửa- Năm 1905, William Bateson, Edith Rebecca Saunders và Reginald Punnett đã phát hiện một số ngoại lệ không tuân theo quy luật Mendel mới được phát hiện lại cách đó không lâu. Trong đó nổi bật là thí nghiệm trên cây đậu thơm: Cây PpLl tự thụ phấn không sinh ra kiểu hình phân li theo tỉ lệ 9:3:3:1, mà lại là xấp xỉ 14:1:1:3. Đây là thí nghiệm đầu tiên đề cập tới tái tổ hợp, nhưng họ đã giả thuyết rằng các gen P/p và L/l là "cặp đôi" với nhau (xem chi tiết ở trang Gen liên kết).
- Sau đó, vào khoảng năm 1910 - 1915, Thomas Hunt Morgan đã phát hiện và giải thích cơ chế đó là do gen hoán vị. Trong quá trình này, đoạn nhiễm sắc thể mang các gen trên cặp nhiễm sắc thể không chị em đã đổi chỗ cho nhau. Nghĩa là đoạn nhiễm sắc thể này chuyển vị sang vị trí tương ứng trên nhiễm sắc thể kia và ngược lại, do đó sự trao đổi "có đi, có lại" giữa hai nhiễm sắc thể tương đồng, tạo ra biến dị "tái tổ hợp" (hình 1).
- Bởi thế, xuất hiện thuật ngữ tái tổ hợp tương đồng để chỉ bản chất tế bào học của hiện tượng này là trao đổi vật chất di truyền (mà mãi sau này mới xác định chính xác là DNA) giữa hai nhiễm sắc thể tương đồng với nhau.
- Vào khoảng cuối những năm 1940, Harriet Creighton và nhất là Barbara McClintock đã chứng minh chi tiết hơn sự trao đổi chéo xảy ra trong quá trình giảm phân ở nhiều loài sinh vật nhân thực.[6]
- Đến năm 1947, nhà vi sinh vật học Joshua Lederberg (hình 2) đã chứng minh vi khuẩn cũng có khả năng giống như sinh sản hữu tính, trong đó có tái tổ hợp gen và đã giành giải Nobel năm 1958.[7]
- Thời gian tiếp theo là những công trình nghiên cứu về tái tổ hợp ở cấp độ phân tử, trong đó sớm nhất là mô hình của Robin Holliday (1964) dựa trên các nghiên cứu của mình về tái tổ hợp ở nấm (Fungi) và mô hình DSBR của Jack Szostak và cộng sự (1983) v.v. Xem chi tiết hơn ở phần dưới.
Đặc điểm chính
sửa- Tái tổ hợp tương đồng (HR) là một kiểu tái tổ hợp gen (cũng gọi là tái tổ hợp di truyền), bao hàm hai sự việc đồng thời:
- Một đoạn DNA ở nhiễm sắc thể này bị cắt, di chuyển sang vị trí tương ứng ở nhiễm sắc thể tương đồng với nó, kết nối vào vị trí đó.
- Cùng lúc, có sự trao đổi ngược lại: một đoạn DNA ở nhiễm sắc thể tương đồng kia cũng bị cắt, di chuyển sang vị trí tương ứng ở nhiễm sắc thể tương đồng với nó, kết nối vào vị trí đó.[8]
- Như vậy, tái tổ hợp tương đồng là một hình thức tái tổ hợp di truyền trong đó các trình tự nu-clê-ô-tit được trao đổi lẫn nhau giữa hai phân tử DNA tương tự nhau hoặc giống hệt nhau.
Vai trò
sửa- HR được các tế bào sống sử dụng rất phổ biến để sửa chữa các sai sót có hại xảy ra trên cả hai mạch đơn DNA khi nhân đôi, theo cơ chế double-strand breaks (viết tắt: DSB, tức là phá vỡ sợi đôi). Quá trình tái tổ hợp tương đồng có thể tạo ra tổ hợp gen mới trong giảm phân, khi hình thành giao tử của sinh vật bậc cao (eukaryote). Nó cũng xảy ra ở vi khuẩn trong quá trình chuyển gen ngang để trao đổi vật liệu di truyền giữa các chủng vi khuẩn. Nhờ đó, tạo điều kiện cho các quần thể có nhiều biến dị thích nghi trong tiến hóa.[1]
- Ngoài vai trò đầu tiên của HR là khả năng phát sinh các alen mới, cung cấp sự đa dạng di truyền giúp chọn lọc tự nhiên hoạt động; thì vai trò thứ hai của HR là được tế bào sử dụng để sửa chữa một số loại tổn thương DNA nhất định giúp cân bằng trong của hệ gen. Thứ ba: ở một số trường hợp ngoại lệ, tái tổ hợp tương đồng là cần thiết cho sự phân hoá thích hợp các nhiễm sắc thể tương đồng trong giảm phân.[9]
Cơ chế tái tổ hợp tương đồng
sửaSinh vật nhân thực
sửa- Sinh vật nhân thực (Eukaryota, cũng gọi là sinh vật nhân chuẩn) có các nhiễm sắc thể tồn tại thành từng cặp tương đồng, trong mỗi cặp thì một chiếc nhận từ bố còn chiếc kia nhận từ mẹ (khác nguồn). Sự tiếp hợp dẫn đến bắt chéo giữa các cặp nhiễm sắc thể tương đồng là cần thiết cho sự phân chia tế bào để hình thành giao tử. Trong quá trình này, tế bào tái tổ hợp tương đồng qua cơ chế hoán vị gen, trong đó sợi đôi DNA bị đứt sẽ được sửa chữa lại để tránh bị sai hỏng trình tự nuclêôtit trong quá trình nhân đôi, dẫn đến sự tái sắp xếp lại các trình tự nuclêôtit đó ở hai nhiễm sắc thể tương đồng khác nguồn.
- Theo những ngiên cứu gần đây, sự tái tổ hợp kết hợp với sửa chữa như thế không phải có thể xảy ra ở bất cứ vị trí nào trên DNA của nhiễm sắc thể, mà chỉ ở những vị trí xác định gọi là điểm nóng tái tổ hợp (recombination hotspots).[10] Sự bắt chéo các nhiễm sắc thể thường bắt đầu chỉ khi có loại prôtêin gọi là Spo11 tạo ra một chỗ đứt trong chuỗi kép DNA đích (cũng gọi là DNA mục tiêu).[11] Thường thì "điểm nóng" này dài khoảng 1.000 - 2.000 bp trên DNA sẽ diễn ra HR.
- Mô hình / con đường tái tổ hợp có khá nhiều, mỗi mô hình/con đường là một giả thuyết xây dựng trên các dữ liệu khoa học, thích hợp với những giai đoạn nhất định trong chu kì tế bào.
- Mô hình sớm nhất là mô hình Holliday (xem ở trang gen hoán vị có giới thiệu sơ lược).
- Mô hình nữa là con đường SDSA (viết tắt từ Synthesis-Dependent Strand Annealing) mô tả có sợi DNA 3 xâm nhập dọc theo 2 sợi đôi DNA nhờ DNA pôlymêraza rồi được giải phóng khi "ngã tư Holliday" hình thành và qua một quá trình được gọi là di chuyển nhánh (hình 3, phải).[12]
- Mô hình DSBR (viết tắt từ Double-Strand Break Repair) bổ sung cho con đường SDSA ở giai đoạn sau khi cắt bỏ, xâm nhập và tổng hợp sợi DNA.[12] (xem giới thiệu sơ lược ở trang gen hoán vị và hình 3, trái).
- Con đường SSA (viết tắt từ Single-Strand Annealing) mô tả sự tái tổ hợp tương đồng có sửa chữa đứt sợi đôi DNA giữa hai trình tự lặp lại (hình 4).
- Con đường BIR (viết tắt từ Break-Induced Replication) là con đường tái tổ hợp tương đồng có sửa chữa các điểm đứt DNA chỉ có một đầu, nghĩa là việc sửa chữa xảy ra ở chạc nhân đôi (chạc chữ Y theo cách dịch ở ta [13]) khi bị hỏng mà vẫn cho phép đầu mút nhiễm sắc thể kéo dài đạt yêu cầu ngay cả khi không có enzym telômeraza.[14]
-
 Hình 2: Joshua Lederberg.
Hình 2: Joshua Lederberg. -
 Hình 3. Phần đầu con đường DSBR và SDSA như nhau, sau thì DSBR thường dẫn đến trao đổi chéo (bên trái), còn SDSA có thể không (bên phải).
Hình 3. Phần đầu con đường DSBR và SDSA như nhau, sau thì DSBR thường dẫn đến trao đổi chéo (bên trái), còn SDSA có thể không (bên phải). -
 Hình 4: Con đường SSA xảy ra giữa hai trình tự lặp lại (màu tím) trên cùng một đoạn DNA dẫn đến mất thông tin di truyền.
Hình 4: Con đường SSA xảy ra giữa hai trình tự lặp lại (màu tím) trên cùng một đoạn DNA dẫn đến mất thông tin di truyền. -
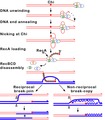 Hình 5: Sơ đồ minh hoạ con đường HR do RecBCD ở vi khuẩn.
Hình 5: Sơ đồ minh hoạ con đường HR do RecBCD ở vi khuẩn.




Ở vi khuẩn
sửa- Vi khuẩn chỉ có 1 DNA-NST (DNA vòng ở vùng nhân), nhưng cũng có HR. Quá trình HR đã được nghiên cứu nhiều nhất và được hiểu rõ nhất ở trực khuẩn lị Escherichia coli. Mặc dù quá trình này khác đáng kể so với tái tổ hợp giảm phân (tức tái tổ hợp tương đồng nhân thực), nhưng nó cũng có trao đổi 2 đoạn DNA, nên cũng gọi là HR.
- HR ở loài vi khuẩn này đã được phát hiện là có ít nhất 25 sản phẩm của gen tham gia, chủ yếu gồm RecA, RecBCD (exonuclease V), RecE (exonuclease VIII), RecF, RecG, RecJ, RecN, Recor, RecQ, RecT, RuvAB, RuvC, SbcCD, và prôtêin SSB (căng mạch), cũng như DNA pôlymêraza I, DNA gyraza, DNA tôpôizômêraza I, DNA ligaza và DNA hêlicaza. Các hoạt động do các sản phẩm này (chủ yếu là enzym) bao gồm ghép cặp DNA tương đồng và trao đổi sợi, tháo xoắn và tách mạch DNA, di chuyển nhánh, liên kết và phân cắt liên kết Holliday, gắn liền các đoạn.[15] Sau đây chỉ giới thiệu về con đường tái tổ hợp RecBCD.
- RecBCD là một phức hợp enzym gồm 3 tiểu đơn vị gọi là RecB, RecC và RecD gắn kết với nhau. Khi phức hợp này di chuyển dọc sợi DNA-NST kép trong lúc nhân đôi, thì nó sẽ khởi tạo HR bằng tháo xoắn DNA và tách đôi mạch khi gặp một chuỗi nuclêôtit gọi là Chi (đọc là "Kai"). Sau đó là 4 giai đoạn sinh hóa: 1 = xử lý phân tử DNA thành các đoạn tái tổ hợp phù hợp; 2 = ghép nối các đoạn DNA thích hợp; 3 = trao đổi sợi DNA và mở rộng DNA; 4 = phân giải cấu trúc bắt chéo.[16]
- RecBCD trượt đến vị trí KAI.
- RecBCD tháo xoắn và cắt hai sợi DNA tạo nên 2 "đuôi" vòng lặp.
- Hai "đuôi" được ủ, tạo ra vòng lặp thứ hai.
- RecBCD tải prôtêin RecA lên đuôi ở vị trí KAI.
- Một đoạn DNA hai mạch còn nguyên vẹn được tải lên, tạo vòng lặp D.
- Vòng lặp D và DNA ban đầu tạo nên "ngă tư" Holliday (cấu trúc 4 sợi bắt chéo).
- Giải tán cấu trúc bắt chéo.
- Xem mô tả con đường RecBCD ở hình 5.
Ứng dụng
sửa- Nhiều phương pháp kĩ thuật đưa đoạn DNA xác định vào sinh vật để tạo ra DNA tái tổ hợp tạo ra các sinh vật biến đổi gen có sử dụng hiểu biết về HR. Các phương pháp này rất phổ biến với đối tượng là nấm men và chuột. Ở chuột, cấc nhà khoa học đã dùng tế bào mầm phôi làm nguồn cấp vật liệu di truyền, kết hợp ức chế gen đích theo nguyên tắc HR. Theo hướng này, các nhà khoa học Mario Capecchi, Martin Evans và Oliver Smithies đã được trao giải Nobel sinh lý học năm 2007.
- Trong di truyền y học, nhiều nhà khoa học cũng đã vận dụng hiểu biết sâu sắc hiện nay về HR để phòng chống một số bệnh ung thư.
Tham khảo thêm
sửaNguồn trích dẫn
sửa- ^ a b “Xuan Li & Wolf-Dietrich Heyer: 'Homologous recombination in DNA repair and DNA damage tolerance'”.
- ^ Campbell: 'Sinh học' - Nhà xuất bản Giáo dục, 2010.
- ^ Max E Gottesman Henry J. Vogel. “Mechanisms of Eukaryotic DNA Recombination”.
- ^ Eric Bank (2018). “Three Mechanisms of Genetic Recombination in Prokaryotes”.
- ^ “Clare O'Connor: 'Meiosis, Genetic Recombination, and Sexual Reproduction'”.
- ^ “Ingrid Lobo & Kenna Shaw: 'Thomas Hunt Morgan, Genetic Recombination and Gene Mapping'”.
- ^ “The Nobel Prize in Physiology or Medicine 1958”.
- ^ “Homologous recombination”.
- ^ “Wayne P. Wahls: Meiotic Recombination Hotspots...”.
- ^ “Meiotic recombination hot spots and cold spots”.
- ^ “Keeney S, Giroux CN, Kleckner N: Meiosis-specific DNA double-strand breaks...”.
- ^ a b “Repair of DNA double-strand breaks by DSBR and SDSA”.
- ^ SGK "Sinh học 12" - Nhà xuất bản Giáo dục, 2016.
- ^ “Break-induced replication: functions and molecular mechanism”.
- ^ S.C Kowalczykowski, D.A Dixon, A.K Eggleston, S.D Lauder & W.M Rehrauer. “Biochemistry of homologous recombination in E.coli”.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Mark S. Dillingham, Stephen C. Kowalczykowski. “RecBCD Enzyme and the Repair of Double-Stranded DNA Breaks”.